Какие ферменты содержатся в цитоплазме гепатоцита
Гепатоциты имеют неправильную многоугольную форму. Средний диаметр клеток — 20-25 мкм. Различают апикальную (билиарную) поверхность гепатоцита, обращенную в просвет желчного капилляра, и базальную (васкулярную) поверхность — в сторону синусоидного капилляра. Своими латеральными поверхностями гепатоцитоты формируют печеночные балки. В центральной части клетки лежит одно-два округлых ядра. Часть из них представляет собой крупные, полиплоидные ядра. Причем число таких ядер увеличивается с возрастом и может достигать в старости 80%.
В цитоплазме хорошо развита гранулярная эндоплазматическая сеть, участвующая в синтезе белков крови. Метаболизм углеводов связан с гладкой эндоплазматической сетью, которая рассеяна в цитоплазме в виде трубочек и пузырьков. Вблизи этих элементов гладкой эндоплазматической сети выявляются гранулы гликогена. Цитоплазма гепатоцитов изобилует митохондриями, число которых в одной клетке превышает 1000. Комплекс Гольджи хорошо развит. Встречаются пероксисомы, лизосомы, а также различные включения (жировые, пигментные и др.).
Количество включений в гепатоцитах находится в связи с фазами пищеварения. После приема пищи резко возрастает количество гликогена, увеличивается число липидных включений. Для печени характерен выраженный суточный ритм: синтез и выделение желчи интенсивнее происходят днем, а гликоген в большом количестве накапливается ночью. Больше гликогена образуется в клетках, расположенных около центральной вены, а желчи — в гепатоцитах на периферии дольки.
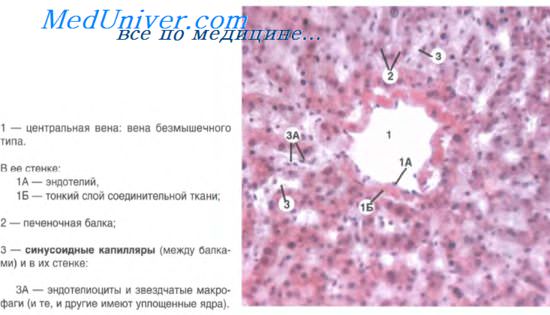
Гепатоциты располагаются обычно в виде двух тесно прилегающих друг к другу рядов, образуя при этом печеночные балки. Между апикальными (билиарными) поверхностями двух гепатоцитов образуется щелевидное пространство с диаметром 0,5-1 мкм. Эти межклеточные узкие щели называют желчными капиллярами. Последние начинаются слепо и в своей начальной части собственной стенки не имеют. Однако ближе к периферии дольки формируются канальцы Геринга — желчные проточки, стенка которых представлена как гепатоцитами, так и эпителиоцитами проточков (холангиоцитами).
По мере увеличения калибра стенка проточка становится сплошной, выстланной однослойным эпителием, в составе которого располагаются малодифференцированные камбиальные холангиоциты. По проточкам желчь попадает в междольковые желчные протоки, выстланные однослойным кубическим эпителием. При обычных методах окраски желчные капилляры не выявляются, но обнаруживаются при импрегнации солями серебра, гистохимической реакцией на щелочную фосфатазу и другими методами.
Таким образом, вырабатывая желчь, печень функционирует как экзокринная железа. Вместе с тем она выделяет в кровь такие вещества, как глюкоза, мочевина, белковые фракции и др., что характеризует печень как эндокринный орган. Из гепатоцитов эти вещества поступают через базальную (синусоидную) поверхность клетки. Между гепатоцитом и гемокапилляром здесь располагается перисинусоидное пространство Диссе, в которое гепатоцит выделяет белки, глюкозу, мочевину и другие вещества в процессе осуществления метаболических функций.
В печеночной дольке существуют две системы, не связанные между собой и действующие по принципу противотока: желчеотводящая, по которой желчь идет от центра на периферию дольки, и кровеносная, по которой кровь движется от периферии к центру дольки. Между желчными и кровеносными капиллярами нет непосредственного соединения, и в условиях нормы желчь не поступает в кровоток. Просвет желчного капилляра является замкнутым благодаря наличию между образующими его соседними гепатоцитами межклеточных контактов нескольких типов — плотных, щелевых и десмосом. В просвет желчного капилляра выступают микроворсинки, образованные на билиарной поверхности гепатоцитов.
Базальная поверхность гепатоцитов обращена в сторону перисинусоидного пространства Диссе. В это пространство выступают также многочисленные микроворсинки, что увеличивает активную поверхность гепатоцитов. Само перисинусоидное пространство, представляет собой узкую щель (шириной 0,2-1 мкм). Если одну стенку его образует базальная поверхность гепатоцитов, то другую — стенка синусоидного гемокапилляра. В пространстве Диссе находятся жидкость, богатая белками, а также аргирофильные фибриллы, единичные фибробласты, отростки звездчатых клеток и др. В нем обнаружены особые мелкие клетки — перисинусоидальные липоциты, или клетки Ито. Они обладают способностью накапливать в цитоплазме липиды и депонировать жирорастворимые витамины. Эти клетки называют также жиронакапливающими, или жирозапасающими, клетками. Их рассматривают как особый тип соединительнотканных интерстициальных клеток.
С функциями клеток связывается синтез и секреция белков коллагена и участие в развитии цирроза печени. В перисинусоидальном пространстве располагаются pit-клетки, относящиеся к большим гранулярным лимфоцитам (натуральные киллеры), которые выделяют вещества, стимулирующие пролиферацию гепатоцитов, участвуют в защитной функции.
Стенка внутридольковых синусоидов выстлана эндотелием, в котором, кроме плоских и тонких эндотелиоцитов, имеются многочисленные вкрапления более крупных звездчатых клеток. Последние известны под названием звездчатые макрофагоциты, или клетки Купфера. Это производные моноцитов крови и представляют собой печеночные макрофаги. В цитоплазме этих клеток много пиноцитоз-ных и фагоцитозных пузырьков, плотных телец (вторичных лизосом). Печеночные макрофаги способны поглощать из крови циркулирующие вещества, накапливать их в цитоплазме, захватывать и переваривать бактерии, обломки эритроцитов. Они способны к амебоидному движению и могут выходить в просвет синусоидов. Набухая, эти клетки выполняют роль сфинктеров синусоидных капилляров.
Эндотелиоциты соединяются в пласт при помощи плотных межклеточных контактов. В выстилке синусоидных капилляров обнаружено наличие мелких отверстий, посредством которых сообщаются между собой просвет синусоидов и пространство Диссе. Поры имеют диаметр около 100 нм. Участки истонченной цитоплазмы эндотелиоцитов, где концентрируются эти отверстия, называют ситовидными пластинками. Они играют роль фильтра. В стенке внутридольковых синусоидных кровеносных капилляров на большом протяжении отсутствует базальная мембрана, что облегчает проникновение веществ из крови в перисинусоидное пространство и в обратном направлении.
Эндотелий синусоидных гемокапилляров, печеночные макрофаги, структуры в перисинусоидном пространстве составляют вместе гепатогематический барьер, или гистион, через который происходит обмен веществ между эпителием печени и кровью.
Наряду с классическими представлениями о строении печеночной дольки, имеются и другие трактовки ее гистоархитектуры. Так, согласно одной из гипотез, элементами дольки являются не печеночные балки, а пластины, состоящие из одного слоя гепатоцитов. Печеночные пластины отгораживают, как стенками, цилиндрические синусоидные пространства (лакуны), по которым протекает кровь.
Кроме классических печеночных долек, описаны так называемые портальные дольки и печеночные ацинусы. Центром портальной дольки признается триада, а периферическими ориентирами являются центральные вены трех смежных долек. В целом портальная долька имеет форму треугольника. В ее пределах кровь течет по направлению от центра на периферию. Печеночный ацинус образуют сегменты двух соседних классических долек, расположенных между близлежащими центральными венами. Ацинус имеет ромбовидную форму. У острых углов ромба находятся центральные вены, а у тупого — триада.
Эти представления о структурно-функциональных единицах печени помогают понять особенности поражений разных отделов печеночной дольки в условиях патологии.
Возрастные изменения печени характеризуются понижением метаболической и пролиферативной активности гепатоцитов, накоплением в их цитоплазме липофусцина и дистрофическими явлениями. Между печеночными дольками разрастается соединительная ткань. Иногда это сопровождается явлениями цирроза печени.
Реактивность и регенерация печени. Ткани печени отличаются высокой чувствительностью к действию повреждающих факторов. Действие ОВ, ионизирующей радиации, комбинированных повреждений приводит к резкому нарушению кровообращения в печени, связанного с его особенностями в этом органе. Нарушается интеграция гепатоцитов в составе печеночных балок, в клетках снижается количество гликогена, изменяется активность окислительно-восстановительных ферментов, подавляется фагоцитарная активность печеночных макрофагов. На месте гибнущих гепатоцитов разрастается рыхлая волокнистая соединительная ткань.
Эпителий печени проявляет способность к физиологической и репаративной регенерации. При удалении в эксперименте на животных до 70% массы печени уже через 2 недели происходит полное восстановление. Этот феномен наблюдается каждый раз при многократных резекциях, проводимых с интервалом около месяца. Однако высокая регенерационная способность печени не характерна для человека. В целом, гепатоциты и холангиоциты относятся к растущей клеточной популяции.
Учебное видео анатомии печени, строения и схемы печеночной дольки

— Вернуться в оглавление раздела «гистология»
Морфофункциональная гепатология
| Сайт: | Образовательный портал МБФ (ВолгГМУ) |
| Курс: | Медицинская биохимия, принципы измерительных технологий в биохимии, патохимия, диагностика, биохимия злокачественного роста. Часть 2. |
| Книга: | Морфофункциональная гепатология |
Оглавление
1. Введение
Наиболее распространено деление органа на правую и левую доли, которые в свою очередь состоят из сегментов. Всего насчитывается 8 сегментов. К каждому из них подходят ветви воротной вены, печеночной артерии, а выходят желчный проток и печеночная вена.
На нижней поверхности печени, примыкающей к воротам, расположена небольшая хвостатая доля (спигелиева). Она соответствует первому сегменту.
В эксперименте удаление части печени ведет к быстрой регенерации этого участка. Так, например, удаление 2/3 печени крысы ведет к восстановлению ее прежних размеров всего за несколько дней. Однако полное восстановление печени ad integrum, по-видимому, возможно при удалении не более 10% массы органа (Bucher N. et al„ 1979).
Установлено, что если хвостатая доля остается неповрежденной, то регенерация печени протекает быстрее (Foucou В. et al., 1983; Soyeux Н. et al., 1984, и др.).
Распределение ферментов в различных отделах печени неодинаково. Так, по данным Е. Schmidt и F. Schmidt (1982), при преимущественном поражении центральных отделов дольки (острый алкогольный гепатит и особенно острый венозный застой) в сыворотке крови нарастает активность глутаматдегидрогеназы, при преимущественном поражении портальных трактов и прилегающих отделов печеночной дольки (острый вирусный гепатит и хронический активный гепатит) в первую очередь повышается активность аминотрансфераз.
2. Гепатоцит
В гепатоците различают васкулярный полюс, в котором при помощи микроворсин осуществляется эндоцитоз (захват веществ извне и введение их в клетку) и экзоцитоз, и билиарный полюс, где при помощи микроворсин происходит выделение веществ из клетки.

Цитоплазматическая мембрана гепатоцита
Отдел мембраны, прилегающий к синусу, занимает 40-50% поверхности всей оболочки. Мембрана высокоактивна в отношении избирательного переноса веществ из крови в гепатоцит, а также из гепатоцита в кровь. Контакт между двумя соседними гепатоцитами осуществляется при помощи латеральной мембраны, занимающей 30-40% поверхности клетки. Большое количество щелей в этой мембране способствует обмену веществ между двумя соседними гепатоцитами.
В сторону желчных ходов обращена канальцевая мембрана, занимающая около 10% поверхности клетки. Мембрана образует большое количество выростов, особенно богатых щелочной фосфатазой, γ-глутамилтрансферазой и 5-нуклеотидазой, получившими вследствие этого название мембранозависимых ферментов. Ферменты обнаруживаются в различных органоидах и отделах клеток. Большинство ферментов локализуется преимущественно в определенных отделах гепатоцита.
Цитоплазма (цитозоль, гиалоплазма) составляет большую часть клетки и содержит различные органоиды, в частности пластинчатый комплекс (тельца Гольджи).
В цитоплазме обнаруживаются следующие ферменты: аланинаминотрансфераза (АлАТ); часть фермента аспартатаминотрансфераза (АсАТ); лактатдегидрогеназа (ЛДГ); сорбит-дегидрогеназа (СДГ); или L-идитолдегидрогеназа; орнитин-карбамоилтрансфераза (ОКТ); часть фермента аргиназа (Apr); часть фермента алкогольдегидрогеназа (АлДГ); γ-глутамилтрансфераза или γ-глутамилтранспептидаза (ГГТФ или ГГТП); фруктозобисфосфат-альдолаза (АЛД), глутатионтрансфераза.
Митохондрии размещены в основном около ядра и в зоне гранулярного эндоплазматического ретикулума у васкулярного полюса гепатоцита. Они производят основную энергию, необходимую для жизнедеятельности клетки. Гепатоцит особенно богат митохондриями; количество их в одной клетке достигает тысячи. В них осуществляются в значительной мере цикл Кребса, окислительное фосфорилирование и окисление жирных кислот. Клинические наблюдения над колебаниями активности глутаматдегидрогеназы дают основание предполагать прямую связь части митохондрий с пребилиарными образованиями клетки.
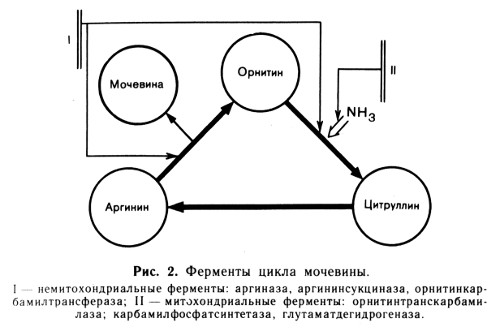
Ферменты, сосредоточенные в основном в митохондриях: часть фермента аспартатаминотрансфераза (АсАТ), глутаматдегидрогеназа (ГлДГ), часть фермента алкогольдегидрогеназа (АлДГ), карбамоилфосфатсинтетаза (КФС), орнитин-транскарбамилаза (ОТК).
В противоположность синдрому Рея при печеночноклеточной коме в основном подавляется активность внемитохондриальных ферментов, участвующих в цикле преобразования мочевины: аргининсукцинатсинтетазы, аргининсукциназы и аргиназы
Гранулярный эндоплазматический ретикулум расположен в основном вокруг ядра и митохондрий. Основу зернистой цитоплазматической сети составляют цистерны, наружная поверхность которых усеяна рибосомами. В них образуются плазмопротеины, за исключением иммуноглобулинов. Синтезируемые белки, в первую очередь альбумины и фибриноген, и большая часть глобулинов через пластинчатый аппарат Гольджи выводятся из гепатоцитов в кровь. Здесь же синтезируются и основные факторы свертывания крови.
Гладкий эндоплазматический ретикулум, или гладкая цитоплазматическая сеть, расположен вокруг ядра и митохондрий и около билиарного полюса гепатоцита. В нем осуществляются биотрансформация и конъюгация как веществ эндогенного происхождения, так и чужеродных для организма соединений (ксенобиотиков). С деятельностью этого органоида в значительной мере связано образование и накопление гликогена. Предполагается, что преимущественно здесь же осуществляется детоксикационная функция печени. В частности, в этих органоидах происходит конъюгация (соединение) токсических веществ с глюкуроновой кислотой, таким путем вещества из жирорастворимых переходят в водорастворимые. Здесь, в частности, осуществляется конъюгация билирубина. Предполагается, что цитохром Р-450, играющий важную роль в преобразовании лекарств и иных ксенобиотиков, в основном сосредоточен в этой фракции. Индикатором Р-450 служит антипириновая проба. Белковая часть глико- и липопротеидов образуется в гранулярном эндоплазматическом ретикулуме и поступает в гладкий эндоплазматический ретикулум, где используется для синтеза этих сложных белков. В этом процессе участвует также пластинчатый комплекс.
В гладком эндоплазматическом ретикулуме обнаруживают глюкозо-6-фосфатазу, билирубингликозилтрансферазу (УДФ-глюкуронилтрансферазу).
Пластинчатый комплекс (аппарат Гольджи) частично с помощью лизосом осуществляет транспорт из клетки ряда веществ, образующихся в эндоплазматическом ретикулуме. Часть этих веществ претерпевает во время транспортировки существенные преобразования. В частности, под влиянием гликозилтрансфераз к молекулам белка присоединяются углеводные комплексы. Таким путем синтезируется часть гликопротеидов или гликозаминогликанов.
Ферменты, сосредоточенные в пластинчатом комплексе, пока в сыворотке крови обычными методами не обнаружены.
Резкие изменения pH среды и повреждения лизосом могут приводить к активации гидролаз и разрушению клетки. Таковы, в частности, элементы патогенеза субмассивных и массивных некрозов печени.
Ферменты, сосредоточенные в основном в лизосомах: кислая фосфатаза, катепсин D, β-глюкуронидаза, β-гексозаминидаза, α-глюкозидаза, рибонуклеаза, дезоксирибонуклеаза.
Микроворсинки билиарного полюса гепатоцита формируют истоки желчных канальцев, играют важную роль в выведении продуктов жизнедеятельности гепатоцитов.
Ферменты, сосредоточенные в основном в микроворсинках: щелочная фосфатаза (ЩФ), 5-нуклеотидаза (5-НТ), аланинаминопептидаза (ААП), γ-глутамилтрансфераза или γ-глутамилтранспептидаза (часть ее сосредоточена в цитоплазме).
Взаимоотношения желчных капилляров (канальцев) и микроворсинок играют важную роль в нормальной секреции желчи. Стенки желчного капилляра образованы канальцевыми отделами мембран гепатоцитов. От места появления эпителия желчного хода и формирования таким путем самостоятельной стенки желчного канала этот отрезок желчевыводящих путей называется мельчайшим желчным протоком.
3. Кле́тки Купфера
Кле́тки Купфера (синонимы: клетки Бровича — Купфера, клетки береговые, клетки синусоидные, клетки эндотелиальные звездчатые) — специализированные макрофаги печени, являющиеся частью ретикулоэндотелиальной системы. Основной функцией клеток Купфера является захват и переработка старых нефункциональных клеток крови. При этом разрушаются молекулы гемоглобина, их глобиновые цепи повторно утилизируются, а гем расщепляется на железо и билирубин.
Названы в честь Карла Вильгельма Купфера, впервые описавшего их в 1876 году
4. Клетки Ито
Клетки Ито (синонимы: звёздчатая клетка печени, жирозапасающая клетка, липоцит, англ. Hepatic Stellate Cell, HSC, Cell of Ito, Ito cell ) — перициты, содержащиеся в перисинусоидальном пространстве печёночной дольки, способные функционировать в двух различных состояниях — спокойном и активированном. Активированные клетки Ито играют главную роль в фиброгенезе — формировании рубцовой ткани при повреждениях печени.
В неповрежденной печени, звёздчатые клетки находятся в спокойном состоянии. В таком состоянии клетки имеют несколько выростов, охватывающих синусоидный капилляр. Другой отличительной чертой клеток является присутствие в их цитоплазме запасов витамина А (ретиноида) в форме жировых капель. Спокойные клетки Ито составляют 5-8 % численности всех клеток печени. [1]
Выросты клеток Ито подразделяются на два типа: перисинусоидальные (субэндотелиальные) и интергепатоцеллюлярные. Первые выходят из тела клетки и простираются вдоль поверхности синусоидного капилляра, охватывая его тонкими пальцеобразными ответвлениями. Перисинусоидальные выросты покрыты короткими ворсинками и имеют характерные длинные микровыбросы, простирающиеся ещё дальше по поверхности эндотелиальной трубки капилляра. Интергепатоцеллюлярные выросты, преодолев пластинку гепатоцитов и достигнув соседнего синусоида, делятся на несколько перисинусоидальных выростов. Таким образом, клетка Ито в среднем охватывает чуть больше двух соседних синусоидов. [2]
При повреждении печени клетки Ито переходят в активированное состояние. Активированный фенотипхарактеризуется пролиферацией, хемотаксисом, сокращаемостью, потерей запасов ретиноида [3] и образованием клеток, напоминающих миофибробластные. Активированные звёздчатые клетки печени также демонстрируют повышенное содержание новых генов, таких как α-SMA, ICAM-1, хемокины и цитокины. Активация свидетельствует о начале ранней стадии фиброгенеза и предшествует повышенному продуцированию ЕСМ-белков. Финальная стадия заживления печени характеризуется усиленным апоптозом активированных клеток Ито, вследствие чего их количество резко сокращается. [4]
Для визуализации клеток Ито при микроскопии применяется окрашивание хлоридом золота. Установлено также, что надёжным маркером для дифференциации этих клеток от других миофибробластов является экспрессия ими белка рилин. [5]
История
В 1876 году Карл Фон Купфер описал клетки, названные им «Sternzellen» (звёздчатые клетки). При окрашивании оксидом золота, в цитоплазме клеток были заметны включения. Ошибочно сочтя их фрагментами эритроцитов, захваченных путём фагоцитоза, Купфер в 1898 году пересмотрел свои взгляды о «звёздчатой клетке» как об отдельном типе клеток и отнес их в разряд фагоцитов. Однако в последующие годы регулярно появлялись описания клеток, похожих на Купферовские «звёздчатые клетки». Им присваивались различные названия: интерстициальные клетки, парасинусоидные клетки, липоциты, перициты. Роль этих клеток оставалась загадкой на протяжении 75 лет, пока профессор Тосио Ито (Toshio Ito) не обнаружил в перисинусоидальном пространстве печени человека некие клетки, содержащие вкрапления жира. Ито назвал их «shibo-sesshu saibo» — жиропоглощающие клетки. Поняв, что вкрапления были жиром, выработанным клетками из гликогена, он сменил название на «shibo-chozo saibo» — жирозапасающие клетки. [6] В 1971 Кендзиро Вакэ (Kenjiro Wake) доказал идентичность «Sternzellen» Купфера и жирозапасающих клеток Ито. Вакэ также установил, что эти клетки выполняют важную роль складирования витамина А (до этого считалось, что витамин А откладывается в клетках Купфера [7] [8] ). Вскоре после этого, Кент и Поппер продемонстрировали тесную связь клеток Ито с фиброзом печени. Эти открытия положили начало процессу детального изучения клеток Ито.

Вверху — схематическое изображение клетки Ито (HSC) по соседству с ближайшими гепатоцитами (PC), ниже синусоидальных эпителиальных клеток печени (EC). S — синусоид печени; KC — клетка Купфера. Внизу слева — клетки Ито в культуре под световым микроскопом. Внизу справа — электронная микроскопия позволяет разглядеть многочисленные жировые вакуоли (L) клеток Ито (HSC), в которых хранятся ретиноиды.
5. Образование желчи
Конъюгированный билирубин секретируется гепатоцитом в желчный капилляр (каналец). Секреция осуществляется в зоне микроворсинок, к мембранам которых очень близко прилегают лизосомы и аппарат Гольджи. По современным воззрениям, желчные канальцы, микроворсинки, прилегающие к ним лизосомы и пластинчатый комплекс составляют желчьсекретирующий аппарат. Этот аппарат секретирует билирубин, холестерин, желчные кислоты, электролиты, воду и другие компоненты желчи. Характерной особенностью желчного канальца является его полная изоляция от кровеносных капилляров.
Тонкий механизм секреции желчи предусматривает выделение холестерина, желчных кислот и пигментов, а также фосфолипидов (в основном лецитина) в виде специфического макромолекулярного комплекса, или желчной мицеллы. Соотношение этих четырех основных компонентов, составляющих мицеллу, в нормальных условиях достаточно постоянно и обеспечивает растворимость компонентов. Ведь холестерин почти нерастворим в воде, плохо растворим в растворе желчных солей, но сочетание желчных кислот и лецитина резко повышает растворимость холестерина. Желчные кислоты, а также хлорид натрия в значительной мере определяют осмотическое давление желчи, а следовательно, и экскрецию воды из гепатоцита.
Знание ультраструктуры гепатоцита и, в том числе, размещения в нем ферментов становится необходимым в клинической практике, в частности в диагностике синдромов и заболеваний печени, в объяснении патогенеза ряда болезней печени (синдром Рея, болезнь Гоше и др.) и в оценке эффективности лечения.
Жёлчь, выделяемая печенью (часть её направляется непосредственно в двенадцатиперстную кишку), называют «печёночной» (или «молодой»), а выделяемую жёлчным пузырём — «пузырной» (или «зрелой»).
Общие характеристики печёночной и пузырной жёлчи [4]
Гепатоцит
В гепатоците различают васкулярный полюс, в котором при помощи микроворсин осуществляется эндоцитоз (захват веществ извне и введение их в клетку) и экзоцитоз, и билиарный полюс, где при помощи микроворсин происходит выделение веществ из клетки.
Цитоплазматическая мембрана гепатоцита
Отдел мембраны, прилегающий к синусу, занимает 40-50% поверхности всей оболочки. Мембрана высокоактивна в отношении избирательного переноса веществ из крови в гепатоцит, а также из гепатоцита в кровь. Контакт между двумя соседними гепатоцитами осуществляется при помощи латеральной мембраны, занимающей 30-40% поверхности клетки. Большое количество щелей в этой мембране способствует обмену веществ между двумя соседними гепатоцитами.
В сторону желчных ходов обращена канальцевая мембрана, занимающая около 10% поверхности клетки. Мембрана образует большое количество выростов, особенно богатых щелочной фосфатазой, γ-глутамилтрансферазой и 5-нуклеотидазой, получившими вследствие этого название мембранозависимых ферментов. Ферменты обнаруживаются в различных органоидах и отделах клеток. Большинство ферментов локализуется преимущественно в определенных отделах гепатоцита.
Цитоплазма (цитозоль, гиалоплазма) составляет большую часть клетки и содержит различные органоиды, в частности пластинчатый комплекс (тельца Гольджи).
В цитоплазме обнаруживаются следующие ферменты: аланинаминотрансфераза (АлАТ); часть фермента аспартатаминотрансфераза (АсАТ); лактатдегидрогеназа (ЛДГ); сорбит-дегидрогеназа (СДГ); или L-идитолдегидрогеназа; орнитин-карбамоилтрансфераза (ОКТ); часть фермента аргиназа (Apr); часть фермента алкогольдегидрогеназа (АлДГ); γ-глутамилтрансфераза или γ-глутамилтранспептидаза (ГГТФ или ГГТП); фруктозобисфосфат-альдолаза (АЛД), глутатионтрансфераза.
Митохондрии размещены в основном около ядра и в зоне гранулярного эндоплазматического ретикулума у васкулярного полюса гепатоцита. Они производят основную энергию, необходимую для жизнедеятельности клетки. Гепатоцит особенно богат митохондриями; количество их в одной клетке достигает тысячи. В них осуществляются в значительной мере цикл Кребса, окислительное фосфорилирование и окисление жирных кислот. Клинические наблюдения над колебаниями активности глутаматдегидрогеназы дают основание предполагать прямую связь части митохондрий с пребилиарными образованиями клетки.
Ферменты, сосредоточенные в основном в митохондриях: часть фермента аспартатаминотрансфераза (АсАТ), глутаматдегидрогеназа (ГлДГ), часть фермента алкогольдегидрогеназа (АлДГ), карбамоилфосфатсинтетаза (КФС), орнитин-транскарбамилаза (ОТК).
В противоположность синдрому Рея при печеночноклеточной коме в основном подавляется активность внемитохондриальных ферментов, участвующих в цикле преобразования мочевины: аргининсукцинатсинтетазы, аргининсукциназы и аргиназы
Гранулярный эндоплазматический ретикулум расположен в основном вокруг ядра и митохондрий. Основу зернистой цитоплазматической сети составляют цистерны, наружная поверхность которых усеяна рибосомами. В них образуются плазмопротеины, за исключением иммуноглобулинов. Синтезируемые белки, в первую очередь альбумины и фибриноген, и большая часть глобулинов через пластинчатый аппарат Гольджи выводятся из гепатоцитов в кровь. Здесь же синтезируются и основные факторы свертывания крови.
Гладкий эндоплазматический ретикулум, или гладкая цитоплазматическая сеть, расположен вокруг ядра и митохондрий и около билиарного полюса гепатоцита. В нем осуществляются биотрансформация и конъюгация как веществ эндогенного происхождения, так и чужеродных для организма соединений (ксенобиотиков). С деятельностью этого органоида в значительной мере связано образование и накопление гликогена. Предполагается, что преимущественно здесь же осуществляется детоксикационная функция печени. В частности, в этих органоидах происходит конъюгация (соединение) токсических веществ с глюкуроновой кислотой, таким путем вещества из жирорастворимых переходят в водорастворимые. Здесь, в частности, осуществляется конъюгация билирубина. Предполагается, что цитохром Р-450, играющий важную роль в преобразовании лекарств и иных ксенобиотиков, в основном сосредоточен в этой фракции. Индикатором Р-450 служит антипириновая проба. Белковая часть глико- и липопротеидов образуется в гранулярном эндоплазматическом ретикулуме и поступает в гладкий эндоплазматический ретикулум, где используется для синтеза этих сложных белков. В этом процессе участвует также пластинчатый комплекс.
В гладком эндоплазматическом ретикулуме обнаруживают глюкозо-6-фосфатазу, билирубингликозилтрансферазу (УДФ-глюкуронилтрансферазу).
Пластинчатый комплекс (аппарат Гольджи) частично с помощью лизосом осуществляет транспорт из клетки ряда веществ, образующихся в эндоплазматическом ретикулуме. Часть этих веществ претерпевает во время транспортировки существенные преобразования. В частности, под влиянием гликозилтрансфераз к молекулам белка присоединяются углеводные комплексы. Таким путем синтезируется часть гликопротеидов или гликозаминогликанов.
Ферменты, сосредоточенные в пластинчатом комплексе, пока в сыворотке крови обычными методами не обнаружены.
Резкие изменения pH среды и повреждения лизосом могут приводить к активации гидролаз и разрушению клетки. Таковы, в частности, элементы патогенеза субмассивных и массивных некрозов печени.
Ферменты, сосредоточенные в основном в лизосомах: кислая фосфатаза, катепсин D, β-глюкуронидаза, β-гексозаминидаза, α-глюкозидаза, рибонуклеаза, дезоксирибонуклеаза.
Микроворсинки билиарного полюса гепатоцита формируют истоки желчных канальцев, играют важную роль в выведении продуктов жизнедеятельности гепатоцитов.
Ферменты, сосредоточенные в основном в микроворсинках: щелочная фосфатаза (ЩФ), 5-нуклеотидаза (5-НТ), аланинаминопептидаза (ААП), γ-глутамилтрансфераза или γ-глутамилтранспептидаза (часть ее сосредоточена в цитоплазме).
Взаимоотношения желчных капилляров (канальцев) и микроворсинок играют важную роль в нормальной секреции желчи. Стенки желчного капилляра образованы канальцевыми отделами мембран гепатоцитов. От места появления эпителия желчного хода и формирования таким путем самостоятельной стенки желчного канала этот отрезок желчевыводящих путей называется мельчайшим желчным протоком.