Что такое кроветворная ткань
При единой концепции кровь рассматривается как мезенхинальная ткань, слагающаяся из клеточного компонента и основного вещества. С точки зрения топографического распространения в организме, структуры и функции, мезенхимальная кровяная ткань состоит из жидкого циркулирующего компонента — периферической крови, и центрального тканевого компонента с более плотной клеточной массой — органов кроветворения. По современным знаниям центральный компонент состоит из двух кроветворных тканей — костного мозга — для эритромиелоидных и мегакариоцитных структур, и лимфоидных органов — для лимфоцитных клеточных структур.
Концепция о единстве крови основывается, в первую очередь на непрерывное взаимоотношение между центральными компонентами, в качестве кроветворных восстанавливающих и дифференцирующих органов, и периферическим отделом, представляющим собой клеточную массу, порождаемую первыми в процессе непрерывной взаимосвязи гомеостатической регуляции. Учитывая тот факт, что периферическая кровь содержит одноядерные (лимфоциты, моноциты) и собственно миелоидные (гранулоциты, эритроциты, тромбоциты) клетки, периферическая кровь представляет собой ткань связи между этими глубоко дифференцированными клеточными линиями с различными функциями.
В их происхождении дифференциация отмечается уже на первых месяцах зачаточной жизни и остается четко отграниченной до конца постэмбрионального периода. Лимфоидные клетки восстанавливаются в лимфатических узлах и прочих лимфоидных структурах (селезенке, миндалинах, пищеварительном тракте), в то время как миелоидные клетки восстанавливаются в костном мозге во время послезародыщевой жизни, спустя определенный период развития на последних 5-6 месяцах зачаточной жизни в печени и селезенке.
Клеточная система периферических макрофагов — моноциты — в классическом понятии зарождаются во всех тканях ретикуло-эндотелиальной структуры (костный мозг, селезенка, лимфоидные органы).
По современной концепции эта система происходит, в основном или даже исключительно, из костного мозга, подобно зернистому ряду (Vaughan, Van Furth). Казалось бы, что в связи с различными функциями и происхождением периферические кровяные клетки не могут быть охвачены единой концепц ей кровяной ткани. Тем не менее большое сходство между существующими планами структурной организации восстановительных органов, равно как и эмбриологическая, онтогенетическая и филогенетическая филиации выступают за единство кровяной ткани. К тому же имеется структурная взаимообусловленность порождающих систем той или иной ткани, также функциональные взаимосвязи, сохраняющие единство кровяной ткани.
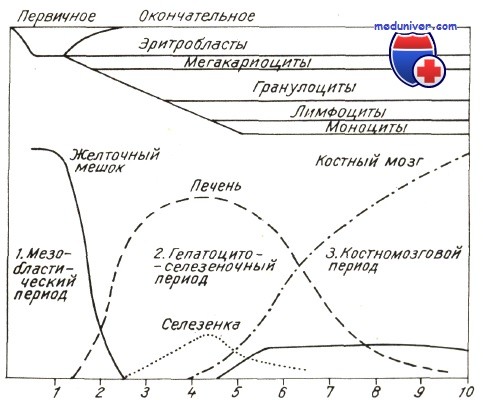 Кроветворение во внутриутробной жизни и на первом месяце послеродового периода
Кроветворение во внутриутробной жизни и на первом месяце послеродового периода
Функциональное и структурное отграничение соответствующих органов является лишь результатом их глубокой дифференциации, однако все они укладываются в широкие рамки кровяной ткани, рассматриваемой как мезенхимальная ткань, общей природы. Как отмечалось ранее, определение крови, по которому последняя рассматривается как единая мезенхимальная ткань, основывается на общие природу и структуру мезенхимальных тканей. Так, все клетки происходят от мезодермального зародышевого листка и состоят из дифференцированных в различной степени клеток и разделенных между собой основным веществом или прочими, происходящими из последнего, мезенхимальными образованиями. Периферические кровяные клетки циркулируют и жизнеспособны в плазме — питательная среда коллоидальной органической структуры, близкой к структуре мезенхимального основного вещества — содержащей комплексные белковые молекулы и агрегаты: мукопротеиды, липопротеиды, мукополисахариды и пр.
Клетки, расположенные в восстановительных органах (костном мозге, лимфоидных органах), раздельно или сгруппированно в колонии и восстановительные единицы, также пропитаны жидким плазматическим основным веществом и разделены, в виде твердой ткани, стромальными, сосудистосинусоидными и фибриллярными структурами. Организационный план восстановительных миелопоэтической и лимфопоэтической тканей общий и сохраняет характеристику первичной дифференцирующейся мезенхимальной ткани. В этом общем организационном плане различаются поддерживающаяся и способствующая трофическим обменам строма и паренхима гемато- или лимфопозтических клеток, обычно организованных в восстанавливающие единицы.
Исходно строма образуется из первичных мезенхимальных клеток, которые, по своей форме, могут быть названы и недифференцированными ретикулярными клетками. При первых попытках объяснить кроветворную филиацию эти клетки были именованы гемогистобластами, спосонбыми размножаться и дифференцироваться в наиболее молодые клетки, восстанавливающие кроветворную активную клеточную систему.
Для дидактических потребностей можно также применять термин гемоцитобласт, определяющий эту промежуточную клетку между недифференцированной ретикулярной в покое и молодой, еще недифференцированной клетками, которая, однако, стремится к восстанавливающей кроветворные клетки эволюции. Нет сомнения, что, в онто- и филогенетическом развитие существует такой план филиации и дифференциации при помощи этой клетки.
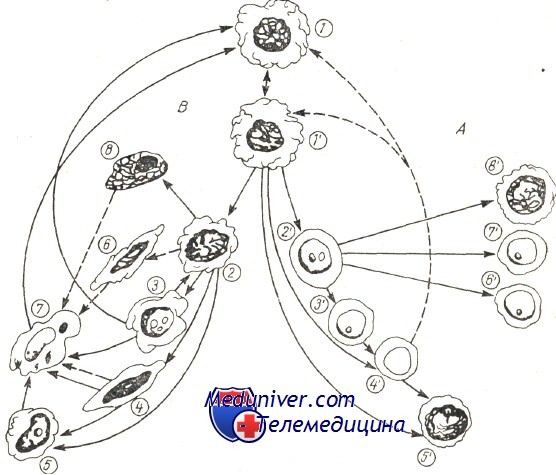 Схема циклической филиации ретикулоэндотелиальных клеток по пути: к образованию гематопоэтических бластических и основных видов ретикулогистиоцитарных клеток.
Схема циклической филиации ретикулоэндотелиальных клеток по пути: к образованию гематопоэтических бластических и основных видов ретикулогистиоцитарных клеток.
Справа (А) изображена филиация процесса кроветворения, а слева (В) филиация клеток ретикулоэндотелиальной системы или системы макрофагов.
Привлекается внимание на обратимость лимфоцита (4′) в гематопоэтическуго клетку-штамм. Обратимость гистиоцита (3) более не допускается;
1) гемоцитобластическая клетка (клетка-штамм или материнская клетка);
2) ретикулярная клетка или гистиобласт;
3) гистиоцит: а) эндотелиальная клетка; б) моноцит; 6) фибробласт; 7) гистиоцит-макрофаг;
8) жировая клетка; 1′) гемогистиобласт; 2′) гемоцитобласт; 3′) лимфобласт; 4′) лимфоцит; 5′) плазмоцит; 6′) миелобласт; 7′) проэритробласт; 8′) мегакариобласт.
Гемоцитобласт — промежуточная клетка — это первая клетка, возглавляющая ряд, восстановливающий все кровяные ряды — лимфоид-ный, эритромиелоцитный, моноцитный, По более старой концепции, под влиянием Ашоффа, вознобленной в отдельных недавно разработанных схемах (Астальди и сотр.), выдвигается идея о том, что в зачаточной и послезачаточной жизни гемоцитобласт, ориентирующийся в направлении кроветворения, берет свое начало от первичной ретикулярной клетки — гемогистиобласта—, как изображено на рисунке, разработанном в монографии «Ретикуло-эндотелиалъная система».
По результатам метода колоний в пробирке или прижизненно из селезенки облученных животных и с пересаженным костным мозгом (Мс Culloch, и Till4) видно, что первичная ретикулярная клетка это видимо неактивная клетка стромы, выполняющая трофическую роль в кроветворении и возможно лишенная какого-либо участия в процессе восстановления крови (Metcalf). Однако по Lajtha и Loutit это восстановительная материнская клетка и в то же время клетка, располагающая трофической активностью — поистине клетка-кормилица, необходимая для восстановления крови.
Кроветворение с образованием бластоклетки, способной восстанавливать все кровяные ряды, в том числе лимфобласт, миелобласт, проэритробласт, мегакариобласт, видимо процесс, берущий свое начало в определенной мезенхимальной клетке, иной чем ретикулярная клетка стромы. Прижизненно эта клетка выявляется при состояниях быстрого восстановления, как, например, в костном мозге после массивного кровотечения или гемолиза, или при патологических состояниях с чрезмерным митотическим делением и блокированием созревания, как бывает при бирмеровской анемии и лейкемиях.
В современных исследованиях образования колоний в пробирке, гемоцитобласт, в качестве общей восстанавливающей кровь клетки, определен, в частности, как клеточная функциональная единица спорной морфологии, получившая недавно — в соответствии с румынской семантикой — следующие названия: клетка зарождения, клетка-штамм, коренная клетка, материнская клетка, что соответствует французскому термину cellule souche, немецкому — Sta-mmzette и особенно весьма распространенному английскому — stem cell. При исследованиях по методу колоний морфологическая характеристика этой клетки соответствует малому лимфоциту, причем обнаруживается она также в периферической крови и в основном в костном мозге. Морфологические критерии дифференциации малого лимфоцита и клетки-штамм еще не уточнены (Rubinstein и Trobpugh).
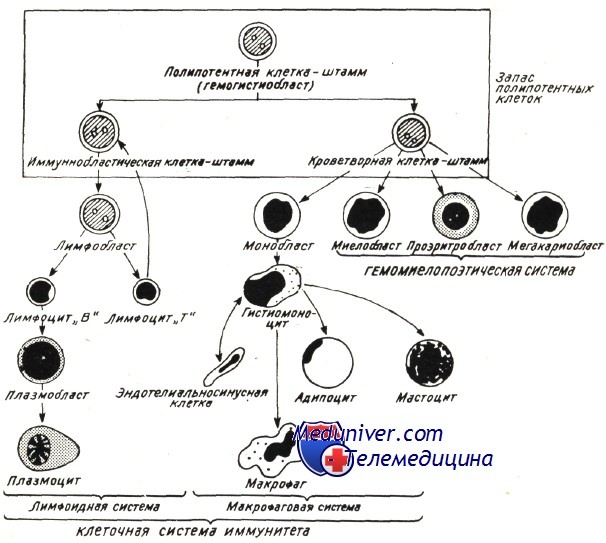 Схема филиации кровяных клеток в рамках дифференцированных систем
Схема филиации кровяных клеток в рамках дифференцированных систем
Методами отстаивания различных градиентов и способами, использующими моноклональные антисыворотки удалось выделить популяцию полипотентных клеток-штамм, равно как и другие популяции клеток-штамм, порождающих остальные кроветворящие линии на стадии созревания.
В эксперименте, после впрыскивания декстенсульфата собаке, Flitner и сотрд. выявили в периферической крови довольно многочисленную популяцию клеток-штамм, наличие которой отмечается в четырех субпопуляциях одноядерных клеток, отделяемых белковым градиентом. У собаки отношение клетки-штамм/одноядерные составляет 1/20 000, в то время как у человека 1/12 000. Вторая фракция не содержит лимфоциты, в то время как клетки-штамм полипотентные, а их структура, исследованая под электронным микроскопом, носит более частный характер, отличаясь от лимфобластов в норме.
Наличие полипотентных клеток-штамм и уже целенаправленных клеток на миело- или лимфоцитные ряды периферической крови и костного мозга также составляют доказательство единства кровяной ткани. К этому следует добавить и общее происхождение в желточном мешке на первом зачаточном периоде. В соответствии с местными структурой и микроклиматом костного мозга или лимфоидной ткани полипотентные клетки-штамм развиваются по типу «целенаправленных » клеток-штамм — возглавляющих ряды дифференцированных линий (Trentin, Fliedner и сотр.).
Восстановление кровяных клеток процессами разрастания и созревания обусловлено гуморальными факторами, такими как эритропоэтин, а. также строма ретикулярных клеток, макрофаги, жировые клетки в трофической взаимосвязи с клетками кроветворной паренхимы.
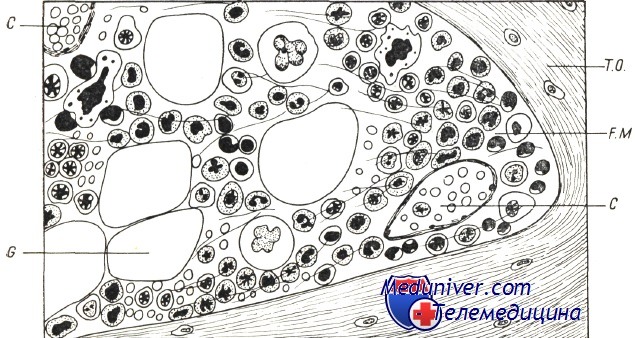 Схема костномозговой структуры в норме.
Схема костномозговой структуры в норме.
С — кровяной капилляр; G—жировое отложение; Т.О.—-костные балки; F.M. — очаг миелопоэза.
В послезачаточной жизни, с точки зрения генетической филиации, исследования методом образования колоний в пробирке или прижизненно показали существование двух, морфологически неразличающихся, видов клетов: 1) коренная клетка, восстанавливающая моно- или полиморфные колонии зернистых миелоидных, эритроцитных, эритробластических, мегакариоцитных и моноцитомакрофаговых клеток; 2) клетка, восстанавливающая лимфоидную линию, порождающую иммунологически компетентные клетки (лимфоцит Т, лимфоцит В и плазмоциты) в условиях нормального иммуного ответа.
Проведенными эмбриологическими исследованиями Максимов первым доказал, что клетка-штамм это лимфоидная клетка, способная восстанавливать все кровяные клетки. Однако современные исследования говорят о том, что, по меньшей мере в после зародышевой жизни, различаются клетки-штамм общей морфологии но различных по функции, при этом одни из них ответственны за миелоидное восстановление в костном мозге, в то время как другие — за лимфоцитное, в лимфоидных органах.
В костном мозге, рассматриваемом как тип кроветворного органа, строма комплекснее, восстанавливающая мозговая ткань располагается в лакунах, образующихся у пересечения костных пластинок губчатых костей. При небольшом разрезе плоской кости с губчатой тканью, например грудины, обнаруживается наличие плотной внутренней и такой же наружной пластинок, между которыми расположена губчатая костная ткань с тонкими пластиночками. Между последними отмечается наличие тонковолокнистой ткани стромы, причем обе они образуют костномозговую структуру стромы.
В состав стромы костного мозга входят кровяные сосуды, артериальные и венозные капилляры, связанные между собой сосудистыми пазухами. Структура последних несложная, она включает прерывистую перегородку из преколлагенных волокон, ретикулиновые волокна, отдельные эластические волокна, на которые опирается прерывистый эндотелий, состоящий из уплощенных первичных ретикулярных клеток.
Между пазухами строму образуют ретикулиновые волокна, пересекающиеся в различных направлениях. У исходного места или в участках пересечения они содержат ретикулярные клетки в покое, амебовидного аспекта, рассматриваемые как гемогистиобласты — первичные мезенхимальные клетки.
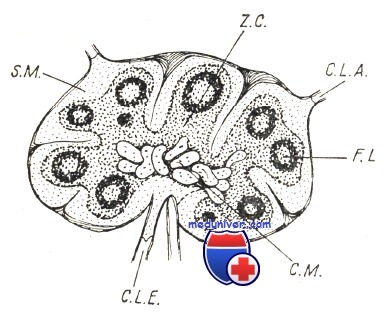 Схема строения лимфатического узла в норме.
Схема строения лимфатического узла в норме.
S.M. — краевой синус; Z.С.—корковая зона; C.L.A. —приводящий лимфатичеческий капилляр; F.L. —лимфатический фолликул; СМ. — костномозговой тяж; C.L.E. — отводящий лимфатический капилляр.
Из этих клеток или из клеток-штамм, зачаточно происходящих от мезенхимальных клеток, рождаются клетки миелоидной ткани в виде мономорфных скоплений эритробластов различного возраста, или миелобластов, разно как и смешанные скопления, в которых возможно наличие мегакариоцитов. В определенных условиях весьма часты скопления эритробластов вокруг ретикулярной клетки, которые, в настоящее время рассматриваются как скопления трофических взаимосвязей между ретикулярной клеткой, носящей характер макрофага и колонией эритробластов. Трофический обмен в этих колониях хорошо изучен посредством переноса железа из макрофаговой ретикулярной клетки к эритробластам на стадии дифференциации (Dacie, и Bessin).
Следовательно, генез колоний не берет свое начало в этой ретикулярной клетке, а, как отмечалось ранее, восстановление осуществляется от лимфоидной клетки-штамм, которая может иметь эритробластическую направленность (Yoffey, Григориу и сотр.). Итак, от клетки-штамм образуются мономорфные колонии определенной направленности или полиморфные колонии, содержащие несколько клеточных видов, в том числе эритробласты, миелобласты, моноциты, мегакариоциты.
Организационный план лимфовосстанавливающих структур близок к костномозговому плану. Также имеется ретикулоклеточная и ретикулосинусоидная стромы и паренхима. В лимфатическом узле нетрудно различить краевой подкапсулярный синус и костномозговые синусоиды. В плотных зонах расположенная балками лимфоидная паренхима и лимфатические фолликулы с зародышевыми центрами маскируют строму. В центре этих зон, после стимула на иммуный ответ (Nossal), становится явной взаимосвязь ретикулярной клетки стромы и подвергающимися лимфобластическому преобразованию лимфоцитами. Как и в отношении миелоидного восстановления, взаимосвязь считается не генетической, а происходящей в порядке трофического обмена, пролиферации и иммуной дифференциации, при этом ретикулярная клетка носит характер не клетки-штамм, а фагоцита.
Располагающиеся вокруг нее лимфоциты образуют «иммуногенные острова», макрофаг, воспринявший антигенный стимул передает специфическую иммуную информацию лимфоцитам Т, которые, в порядке «иммунологического сотрудничества» определяют специфический первичный или вторичный иммуный ответ (Берчану).
Функции этих двух организационных планов обоих схожих систем восстановления клеточных компонентов периферической крови связываются ретикулярной клеткой в процессе макрофаговой активности. И если в настоящее время отвергается идея общего происхождения этих двух систем от ретикулярной клетки, располагающей генетической восстановительной способностью, тем не менее признается ее общая роль в качестве стромальной клетки, выполняющей трофическую функцию стимулирования восстановления и дифференциации.
Четко определена функция макрофага в иммунологическом ответе в рамках иммуного клеточного сотрудничества. Также доказана его функция в процессе восстановления колоний миелоидных клеток (Metcalf), причина по которой он получил название «клетка-кормилца», т.е. питающей клетки. В клеточных культурах, как при бластических преобразованиях лимфоцитов под влиянием РНА, так и при методе образования миелопоэтических колоний доказана в пробирке стимулирующая рост роль макрофагов. В случае отсутствия таковых, развитие колоний либо скудное, либо совсем не отмечается, в то время как на слое макрофагов или фибробластов, плотно прилегающих к стеклу и тем самым образующих однослойную питательную среду, названную весьма выразительно «feeder-layer», процесс развития протекает отлично (Берчану, Гочу и Моцою).
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Что такое кроветворная ткань
Эмбриональный источник развития кроветворных тканей — мезенхима. В эмбриогенезе можно различать 3 периода гемопоэза: 1) внезародышевый, или мезобластический (1-2 мес), 2) гепато-тимо-лиенальный (2-5 мес), 3) медулло-тимо-лимфоидный (5-10 мес).
Постэмбриональным гемопоэзом, или физиологической регенерацией крови, называют кроветворение во взрослом организме.
Кроветворение в желточном мешке. В конце 2-й — начале 3-й недели эмбриогенеза в мезенхиме стенки желточного мешка образуются кровяные островки, в составе которых клетки дифференцируются на плоские эндотелиальные и округлые клетки. Последние преобразуются в стволовые кроветворные клетки. При внезародышевом кроветворении из стволовых клеток формируются первичные эритробласты — мегалобласты. Они делятся внутри сосудистого русла (интраваскулярно). Небольшая часть мегалобластов превращается в безъядерные первичные эритроциты — мегалоциты. Образуется также незначительное количество вторичных эритроцитов меньшей величины, чем мегалоциты. Экстраваскулярно дифференцируется часть первичных лейкоцитов (гранулоцитов — нейтрофилов и эозинофилов). Из желточного мешка стволовые кроветворные клетки по развивающимся сосудам расселяются по органам зародыша.
Кроветворение в печени. На 2-м месяце эмбриогенеза печень становится центром кроветворения. Источником гемопоэза здесь служат стволовые кроветворные клетки. Кроветворение в печени происходит экстраваскулярно. Из стволовых кроветворных клеток образуются эритроциты, зернистые гранулоциты (нейтрофилы и эозинофилы) и мегакариоциты. Зернистые лейкоциты развиваются здесь укороченным путем и не имеют четкой специфической зернистости. К концу эмбриогенеза человека кроветворение в печени постепенно прекращается.
Кроветворение в селезенке. На 4—5-м месяцах эмбриогенеза человека селезенка становится универсальным органом гемопоэза, в котором экстраваскулярно образуются все клетки крови. Позднее процессы эритро- и гранулоцитопоэза в селезенке угасают, но усиливается образование незернистых лейкоцитов.
Кроветворение в красном костном мозге и тимусе. Постепенно центральным органом кроветворения становится красный костный мозг. Строму его вначале образует мезенхима, которая позднее преобразуется в ретикулярную ткань. Ретикулярная ткань, в трехмерной сети которой происходит развитие эритроцитов, гранулоцитов, моноцитов и мегакариоцитов, называют миелоидной тканью. Миелоидная ткань — специализированная гемопоэтическая ткань красного костного мозга. Она обеспечивает развитие стволовых клеток и всех форменных элементов крови. Наряду с миелоидной к кроветворным тканям относится лимфоидная ткань, которая развивается в лимфатических узлах, селезенке и других лимфоидных органах, составляющих лимфоидную систему. Здесь в сети ретикулярной ткани происходит образование лимфоцитов, плазматических клеток, удаление клеток и продуктов их распада.
К центральным органам кроветворения относится тимус, в котором на 2-м месяце эмбриогенеза начинают дифференцироваться лимфоциты тимуса. В дальнейшем они расселяются по периферическим органам лимфоидной системы.
Кроветворение в лимфатических узлах начинается с 4-го месяца эмбриогенеза после миграции стволовых кроветворных клеток.
В соответствии с унитарной теорией кроветворения А.А. Максимова, существует единый источник развития для всех клеток крови. Исходной клеткой для всех ростков кроветворения является стволовая кроветворная клетка, сходная по своему строению с малым лимфоцитом. А.А. Максимов (1911) писал, что индифферентные блуждающие клетки, или лимфоциты в широком смысле, одарены очень большой потенцией развития: «Это индифферентная мезенхимная блуждающая клетка, лимфоцит, является общей родоначальницей всех элементов крови. Попадая в благоприятные условия, она проявляет свою потенцию развития, причем в зависимости от условий, направление развития и продукты его получаются очень разнообразными». Унитарная теория кроветворения была развита в трудах А.А. Заварзина, Н.Г. Хлопина, А.Н. Крюкова, М.И. Аринкина и др. Метод селезеночных колоний, разработанный канадскими учеными Тиллом и МакКуллохом (1961), прозволил идентифицировать вид клеток, являющийся источником развития клеток эритроидного, гранулоцитарного и мегакариоцитарного рядов. Эту клетку, которая гистологически сходна с малым темным лимфоцитом, авторы назвали колониеобразующей единицей (КОЕ).
В развитии клеток крови условно выделяются классы клеток. По мере перехода клеток из класса в класс, в каждом из них все более отчетливо обнаруживаются гемопоэтические клеточные диффероны, которые характеризуются определенными гистологическими признаками. Однако клетки первых трех классов по своему строению идентичны. Только методы иммуноцитохимии позволяют различать клетки по наборам клеточных рецепторов, что является показателем дивергентной дифференцировки стволовой клетки.
В общем виде развитие клеток крови происходит в следующей последовательности.
1-й класс — плюрипотентные клетки — это стволовые кроветворные клетки (СКК). Стволовая клетка является общим самоподдерживающимся предшественником всех клеток крови, включая все виды иммунокомпетентных клеток. Полагают, что каждая из стволовых клеток способна проделать по меньшей мере 100 митозов, т. е. потомками одной стволовой клетки можно было бы обеспечить всю кроветворную систему. Однако стволовые клетки после цикла пролиферации в эмбриогенезе переходят в состояние покоя. Она лишена каких-либо специфических признаков строения и локализуется в миелоидной ткани среди популяции лимфоците- или моноцитоподобных элементов. Она может с током крови мигрировать по тканям организма. Объективным методом обнаружения и количественного учета стволовых клеток является метод селезеночных колоний. Стволовые клетки составляют около 0,1% популяции кроветворных элементов.
2-й класс. Стволовые клетки под влиянием ряда факторов (тромбопоэтический, ИЛ-7 и др.) дивергентно дифференцируются в двух направлениях: полустволовые, или мультипотентные, клетки — предшественники миелопоэза (КОЕ-ГЭММ) и мультипотентные клетки — предшественники лимфопоэза (КОЕ-Л). В составе колоний эти клетки имеют ограниченные возможности к самоподдержанию (около 3-4 недель), однако этого достаточно для поддержания физиологической регенерации крови.
3-й класс. Из мультипотентных клеток — предшественников миелопоэза (КОЕ-ГЭММ) в результате дивергентной дифференцировки, происходящей под влиянием ряда факторов микроокружения, возникают следующие клеточные линии: а) родоначальные (прогениторные) клетки, или клетки-предшественники, эритропоэза (БОЭ-Э, от англ. burst — взрыв) и развивающиеся из них КОЕ-Э; б) общие родоначальные клетки гранулоцито- и моноцитопоэза (КОЕ-ГМо). Последние в процессе дальнейшей дивергентной дифференцировки под влиянием факторов микроокружения формируют родоначальные клетки для нейтрофильных (гранулоцит-стимулирующий фактор), эозинофильных (ИЛ-5) и базофильных (ИЛ-3) гранулоцитов (КОЕ-Гн, КОЕ-Эо, КОЕ-Б) и моноцитов (КОЕ-Мо, фактор — моноцит-колониестимулирующий).
Мультипотентные клетки лимфопоэза (КОЕ-Л) под влиянием дифференцировочных факторов микроокружения (ИЛ-7, ИЛ-6) развиваются в родоначальные клетки Т- и В-лимфоцитов.
Мультипотентные клетки КОЕ-ГЭММ (при участии тромбопоэтина и ИЛ-11) являются источником развития родоначальной клетки для мегакариоцитов (КОЕ-Мег).
Таким образом, важнейшее свойство, которое приобретают в миелопоэзе и лимфопоэзе кровеобразующие клетки — это формирование рецепторно-трансдукторной системы, реагирующей на конкретные факторы дифференцировки (эритропоэтин, тромбопоэтин, колониестимулирующие факторы, интерлейкины — ИЛ и др.), вырабатываемые кроветворным микроокружением и клетками других органов. Все это приводит к тому, что в клетках появляются гистологические маркеры, на основе которых можно с большой вероятностью отнести ту или иную клетку к конкретному гемопоэтическому ряду (дифферону).
IV-й класс клеток — гистологически распознаваемые клетки кроветворной ткани — это пролиферирующие клетки («бласты»). Они способны к пролиферации и дифференцировке.
V-й класс — созревающие клетки («про-циты») и VI класс — зрелые клетки периферической крови.