Ресинтез АТФ в мышечных волокнах
Дано определение ресинтеза АТФ. Описаны основные пути ресинтеза АТФ в мышечных волокнах: креатинфосфатный, гликолитический, миокиназный и тканевое дыхание. Описаны количественные критерии путей ресинтеза АТФ, соотношение между различными путями ресинтеза АТФ при мышечной работе, а также между путями ресинтеза АТФ и зонами относительной мощности.
Ресинтез АТФ в мышечных волокнах
Определение
Ресинтез АТФ – синтез АТФ из различных энергетических субстратов во время физической работы в мышечных волокнах.
Формула ресинтеза АТФ выглядит следующим образом:
Пути ресинтеза АТФ
Ресинтез АТФ может осуществляться двумя путями:
Аэробный путь (тканевое дыхание, аэробное или окислительное фосфорилирование) – основной способ образования АТФ в мышечных волокнах. Он протекает в митохондриях мышечных волокон. В результате тканевого дыхания выделяется 39 молекул АТФ. Окисляемое вещество распадается до углекислого газа и воды.
Анаэробный ресинтез АТФ
Анаэробные пути ресинтеза АТФ являются дополнительными способами образования АТФ в мышечных волокнах в тех случаях, когда основной путь получения АТФ – тканевое дыхание не может обеспечить мышечную деятельность необходимым количество кислорода. Эти механизмы ресинтеза АТФ активно функционируют в начале выполнения физических упражнений, когда тканевое дыхание не полностью «развернулось», а также при физических нагрузках высокой мощности.
Анаэробный ресинтез АТФ в мышечных волокнах возможен посредством нескольких механизмов:
Количественные критерии путей ресинтеза АТФ
Существуют количественные критерии путей ресинтеза АТФ. К ним можно отнести: максимальную мощность, время развертывания, время сохранения или поддержания максимальной мощности, метаболическую ёмкость (табл. 1).
Таблица 1. Количественные критерии основных путей ресинтеза АТФ (С.С. Михайлов, 2009)
| Пути ресинтеза АТФ | Критерии | |||
| Максимальная мощность, кал/мин кг | Время развертывания | Время сохранения максимальной мощности | Метаболическая ёмкость | |
| Креатинфосфатный | 900-1100 | 1-2 с | 8-10 с | |
| Гликолитический | 750-850 | 20-30 с | 2-3 мин. | При анаэробном окислении гликогена образуются 3 молекулы АТФ в расчете на одну молекулу глюкозы |
| Аэробный | 350-450 | 3-4 мин. | Десятки минут | При аэробном окислении гликогена образуются 39 молекул АТФ в расчете на одну молекулу глюкозы (самый экономичный) |
Соотношение между различными путями ресинтеза АТФ
При любой мышечной работе функционируют все три основных механизма ресинтеза АТФ, но включаются они последовательно. В первые секунды ресинтез АТФ осуществляется за счет креатинфосфатной реакции, затем включается гликолиз. По мере продолжения работы на смену гликолизу приходит тканевое дыхание (рис.1). Эта смена механизмов ресинтеза АТФ приводит к уменьшению суммарной выработки АТФ.
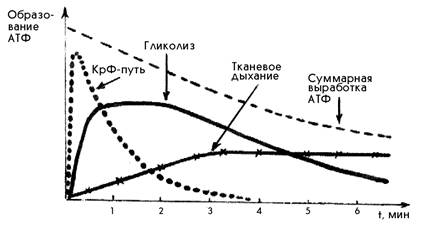
Рис.1. Включение путей ресинтеза АТФ при выполнении физической работы (С.С. Михайлов, 2009)
Пути ресинтеза АТФ и зоны относительной мощности
В.С. Фарфель приводит следующее соотношение мощности работы и основной системы энергообеспечения (табл.2)
Таблица 2. Зоны мощности работы и основная система энергообеспечения (В.С. Фарфель)
Энергетические процессы в мышце
Содержание
Энергетические процессы в мышце [ править | править код ]
Естественно, что для совершения мышечного движения требуется энергия. В организме человека существуют разные источники энергии, которые последовательно включаются один за другим. Рассмотрим каждый из них.
АТФ [ править | править код ]
Универсальным источником энергии в живом организме является молекула АТФ, которая образуется в цитратном цикле Кребса. Под действием фермента АТФазы молекула АТФ гидролизуется, отсоединяя фосфатную группу в виде ортофосфорной кислоты (Н3РО4), и превращается в АДФ, при этом высвобождается энергия.
Головка миозинового мостика при контакте с актином обладает АТФазной активностью и соответственно возможностью расщеплять АТФ и получать энергию, необходимую для движения.
 Количества АТФ, которое содержится в мышце, достаточно для выполнения движений в течение 2-5 первых секунд.
Количества АТФ, которое содержится в мышце, достаточно для выполнения движений в течение 2-5 первых секунд.
Креатинфосфат [ править | править код ]
Запас молекул АТФ в мышце ограничен, поэтому расход энергии при работе мышцы требует постоянного его восполнения, это происходит за счет креатинфосфата. Креатинфосфат обладает способностью отсоединять фосфатную группу и превращаться в креатин, присоединяя фосфатную группу к АДФ, которая превращается в АТФ.
АДФ + креатинфосфат = АТФ + креатин.
Эта реакция получила название – реакции Ломана. Именно поэтому креатин имеет большое значение в бодибилдинге.
Надо заметить, что креатин эффективен только при выполнении анаэробных (силовых) упражнений, так как креатинфосфата достаточно примерно на 2 минуты интенсивной работы, затем подключаются другие источники энергии. Соответственно, в лёгкой атлетике приём креатина как добавки для увеличения атлетических показателей малоэффективен.
Запасы креатинфосфата в волокне не велики, поэтому он используется в качестве источника энергии только на начальном этапе работы мышцы, до момента активизации других более мощных источников – анаэробного и затем аэробного гликолиза. По окончании работы мышцы реакция Ломана идет в обратном направлении, и запасы креатинфосфата в течение нескольких минут восстанавливаются.
Энергетический метаболизм скелетных мышц [ править | править код ]
Алактатные механизмы [ править | править код ]
КФ обеспечивает запас энергии фосфата для ресинтеза АТФ из АДФ при наступлении сократительной деятельности (рис. 3):
КФ + АДФ Креатинкиназа К + АТФ (1)
В состоянии покоя мышечные волокна наращивают концентрацию КФ до пяти раз больше, чем АТФ. В начале сокращения, когда концентрация АТФ начинает падать, а АДФ повышаться вследствие ускорения разложения АТФ, массовая активность способствует образованию АТФ из КФ.
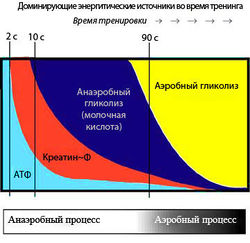
Хотя образование АТФ из КФ происходит быстро, требуя одной единственной ферментативной реакции (1), количество АТФ, которое может быть получено в результате этого процесса, ограничено начальной концентрацией КФ. Мышечные волокна также содержат миокиназу, которая катализирует образование одной молекулы АТФ и одной молекулы АМФ из двух молекул АДФ. АТФ и КФ, вместе взятые, могут обеспечить максимальную силу в течение 8—10 с. Таким образом, энергия, полученная от фосфагенной системы, используется для коротких всплесков максимальной мышечной активности, необходимых в легкой и тяжелой атлетике (забег на 100 м, толкание ядра или поднятие тяжестей).
Гликолиз [ править | править код ]
Хотя метаболизм по гликолитическому пути производит лишь небольшое количество АТФ из каждой усвоенной единицы глюкозы, он может обеспечить быстрый синтез большого количества АТФ при наличии достаточного количества ферментов и субстрата. Этот процесс может также происходить в отсутствие кислорода:
Глюкоза анаэробный быстрый гликолиз 2 АТФ + 2 лактата (2)
Глюкоза для гликолиза поступает либо из крови, либо из запасов гликогена. Когда исходным материалом выступает гликоген, из одной единицы потребленной глюкозы в результате фосфоролитического гликогенолиза образуется три молекулы АТФ. По мере того, как мышечная активность становится интенсивнее, для анаэробного расщепления гликогена мышц требуется все больше и больше АТФ, и, соответственно, увеличивается производство молочной кислоты. Анаэробный гликолиз может обеспечить энергию на 1,3-1,6 мин максимальной мышечной активности.
Образование молочной кислоты понижает уровень pH в мышечных волокнах. Это препятствует действию ферментов и вызывает боль, если удаление молочной кислоты происходит слишком медленно по сравнению с ее образованием.
Окислительное фосфорилирование [ править | править код ]
При умеренном уровне физической нагрузки, например, при беге на 5000 м или марафоне, большая часть АТФ, используемого для сокращения мышц, образуется путем окислительного фосфорилирования. Окислительное фосфорилирование позволяет высвободить из глюкозы гораздо больше энергии по сравнению с отдельно взятым анаэробным гликолизом:
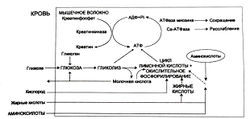
Жиры катаболизируются только с помощью окислительных механизмов, при этом выделяется много энергии. Аминокислоты тоже могут быть метаболизированы подобным образом. Три метаболических пути образования АТФ для сокращения и расслабления мышц показаны на рис. 3.
В течение первых 5
10 мин умеренной физической нагрузки главным потребляемым «топливом» является собственный гликоген мышц. В течение следующих 30 мин доминирующими становятся переносимые кровью вещества; глюкоза крови и жирные кислоты вносят примерно одинаковый вклад в потребление мышцами кислорода. По истечении этого периода все более важную роль приобретают жирные кислоты. Важно подчеркнуть взаимодействие между анаэробными и аэробными механизмами в образовании АТФ во время физической нагрузки. Вклад анаэробного образования АТФ больше при краткосрочной нагрузке высокой интенсивности, в то время как при более продолжительных нагрузках низкой интенсивности преобладает аэробный метаболизм.
Восстановление и кислородная задолженность [ править | править код ]
После того как физическая нагрузка закончилась, поглощение кислорода все еще остается выше нормы (табл.). С недавнего времени для обозначения кислородной задолженности используется также термин «избыточное потребление кислорода после физической нагрузки». Сначала его уровень очень высок, пока тело восстанавливает запасы КФ и АТФ, возвращая тканям запасенный кислород, а затем в течение еще одного часа потребление идет на более низком уровне, пока удаляется молочная кислота. Поэтому ранние и последние фазы кислородной задолженности называют соответственно алактатной и лактатной кислородной задолженностью. Повышение температуры тела также говорит о более высокой скорости метаболизма и росте потребления кислорода.
Чем продолжительнее и интенсивнее физическая нагрузка, тем больше времени занимает восстановление. Например, на восстановление после полного истощения гликогена мышц зачастую требуется несколько дней, а не секунд, минут или часов, необходимых для восстановления запасов КФ и АТФ и удаления молочной кислоты. Физическая нагрузка большой интенсивности, вероятно, приводит к микротравмам мышечных волокон, и их восстановление занимает некоторое время.
Компоненты кислородной задолженности. После длительной, тяжелой физической нагрузки дыхание остается выше нормы для удовлетворения повышенной потребности в кислороде
Восстановление запасов кислорода в тканях(около 1 л)
Восстановление уровней креатинфосфата и других богатых энергией фосфатов (около 1-1,5 л)
Удаление молочной кислоты путем глюконеогенеза и другими путями (до 12 л)
Стимуляция метаболизма вследствие повышения уровня адреналина (около 1 л)
Дополнительное потребление кислорода в дыхательных мышцах и сердце (около 0,5 л)
Общее усиление метаболизма вследствие более высокой температуры тела*
Роль аэробного механизма энергообеспечения в борьбе
В. Н. Селуянов
ПНИЛ, РГАФК, Москва
Соревновательная деятельность в борьбе продолжается мин. и завершается, как правило, предельным утомлением спортсмена. В циклических видах спорта такая деятельность может быть сопоставлена с соревновательной деятельностью бегуна на 1500–3000 м. Поскольку этот вид деятельности хорошо изучен в физиологии, то достаточно легко найти экспериментальные данные и физиологические механизмы, объясняющие результативность такой деятельности.
Механизм энергообеспечения такой работы может быть описан следующим образом. В начале, для преодоления внешнего сопротивления, которое составляет около 40 % от МАМ, должны быть рекрутировано около 40 % МВ. Эти мышечные волокна являются окислительными. В них начинается трата молекул АТФ и ресинтез их за счет энергии молекул КрФ. Свободные Кр и неорганический фосфат активизируют деятельность гликолиза и окисления жиров одновременно. Через после начала упражнения запасы АТФ и КрФ в рекрутированных МВ значительно снижаются, поэтому мощность работы этих мышечных волокон падает в 2–3 раза. Это заставляет спортсмена рекрутировать новые МВ в количестве, необходимом для поддержания заданной мощности. Следовательно, следующие 10–15 с работа поддерживается за счет аэробных процессов в ранее рекрутированных МВ и энергии АТФ и КрФ в новых МВ. Затем, описанный механизм рекрутирования МВ, продолжает развиваться. Начинают подключаться к работе гликолитические МВ, которые после исчерпания запасов АТФ и КрФ начинают работать в анаэробном гликолизе с образованием лактата и ионов водорода. По данным B. Saltin et al. (1972) интенсивный рост концентрации лактата в мышце начинается после минуты работы. Продолжительность работы рекрутированных гликолитических МВ не превышает одной минуты, поскольку закисление МВ приводит к потере силы и мощности их функционирования. Поэтому работа с заданной мощностью будет продолжаться до тех пор, пока есть что рекрутировать. В момент исчерпания всех МВ заданная мощность уже не может больше поддерживаться. В этот момент мышцы предельно закисляются, потребление кислорода, ЧСС и легочная вентиляция достигаю также предельных величин. Испытуемый испытывает тяжелое физиологическое состояние и отказывается от продолжения работы. Если работа продолжалась 6 мин., то за это время потребление кислорода мышцами составит: V(О2) АнП × 6 мин. = 4 л/мин х 6 мин. = 24 л/мин. Если мощность работы составила 400 Вт или 5,3 л О2/мин, то кислородный запрос упражнения составит 5,3 × 6 мин. = 32 л О2. Дефицит кислорода составил 8 л, из него приходится на АТФ и КрФ, а на анаэробный гликолиз 5–6 л.
Следовательно, основным механизмом энергообеспечения является аэробный (24/32 × 100 % = 75 %, алактатный 2/32 × 100 % = 6,3 %, анаэробный гликолиз 6/32 × 100 % = 18,7 %). Эта оценка в целом согласуется с данными многих авторов.
Заметим также, что с ростом потребления кислорода на уровне АнП, когда он приближется по своей величине к МПК, наблюдается увеличение продолжительности работы на уровне МПК, снижается степень закисления мышц и крови. В целом вклад в запрос кислорода аэробных процессов растет и может достигать 90 %.
Отсюда следует приоритет в развитии аэробного механизма энергообеспечения у спортсменов, выполняющих предельную мышечную работу в пределах
Единственным специалистом, результаты которого вошли в противоречие с общеустановленными представлениями является В. В. Шиян (1997). На основании исследования соревновательной деятельности различных видов борьбы он пришел к выводу о приоритете анаэробного механизма энергообеспечения.
На основе анализа биоэнергетических характеристик у спортсменов различных видов спорта им было установлено, что «у борцов существенно меньше показатели бионергетических функций, чем у представителей других видов спорта». Например, борцы имеют аэробную мощность (МПК) 58 мл/мин/кг, тогда как у бегунов на дистанцию 800 м и более длинные более 70 мл/мин/кг. Автор предположил, что отставание в развитии борцов связано с отставанием в теории и методике подготовки высококвалифицированных борцов по сравнению с другими видами спорта.
Эта аргументация поверхностная, а обнаруженное явление легко объяснить, если придерживаться концепции о периферическом лимитирующем факторе МПК. В этом случае потребление кислорода определяется массой митохондрий потребляющих кислород в активных мышцах ног, сердце и дыхательных мышц. При педалировании на велоэргометре основную работу выполняют только мышцы ног, поэтому при делении на массу тела, в которую входят у борцов существенно гипертрофированные и массивные мышцы спины, живота и рук, при равных абсолютных величинах МПК относительные величины МПК у борцов окажутся ниже при сравнении с представителями других видов спорта без существенной гипертрофии мышц пояса верхних конечностей.
Другим аргументом В. В. Шияна стали данные факторного анализа, из которых следовало, что вклад в общую дисперсию аэробных способностей борцов составил только 10 %, а основная дисперсия пришлась на анаэробные возможности — более 90 %. Следовательно, «подготовка борца высокой квалификации (особенно этап подготовки к соревнованиям), должна быть направлена на максимальное развитие преимущественно анаэробноых возможностей спортсменов». Такая аргументация не выдерживает следующей критики. При изучении однородных выборок спортсменов наиболее важные для достижения высоких спортивных результатов показатели у всех спортсменов должны быть примерно равными, следовательно, должны мало варьировать. Показатели, которые существенно варьируют, не имеют принципиального значения для данного вида спорта. Из этого следует, что именно анаэробные показатели не имеют принципиального значения при оценке уровня подготовленности борцов высокой квалификации.
Подтверждение нашей аргументации можно найти при анализе соревновательной деятельности борцов. Например, по данным В. В. Шияна (1997) активность победителей и надежность технических действий выше чем у побежденных на а закисление у победителей либо меньше, либо статистически достоверно не различались (рН = 7,158, б = 0,077). Из этого следует, что более высокая активность борцов победителей определялась их более высокой аэробной подготовленностью.
Другим важным аргументом в пользу важности аэробной подготовленности борцов являются данные о тестировании и норме специальной выносливости борцов. В. В. Шиян (1997) использовал в своих исследованиях тест — педалирование на велоэргометре, три раза по одной минуте предельной нагрузки. После тестирования на мин. бралась кровь из мочки уха, для определения рН. По данным тестирования по формуле определяли гликолитическую эффективность:
АнГЭ = ΣАi/(100 х ΔpH)
, где ΣАi сумма трех работ, выполненных в одноминутных предельных упражнениях, ΔpH — изменение степени закисления крови по данным анализа крови до и после тестирования.
Аналогичная формула использовалась для оценки специальной выносливости (коэффициент специальной выносливости):
Σti — сумма трех работ, выполненных в виде серий по 15 бросков чучела, с.
Анализ этих уравнений показывает, что при равном объеме выполненной работы уменьшение степени закисления крови ведет к росту специальной выносливости. Следовательно, показатели АнГЭ и КСВ характеризуют уровень аэробных возможностей спортсмена. К сожалению результаты, получаемые по этим формулам, получили некорректную интерпретацию. В. В. Шиян (1997) предположил, что одноминутная работа связана преимущественно с анаэробным механизмом энергообеспечения — анаэробным гликолизом, поэтому показатель КСВ должен характеризовать анаэробную гликолитическую мощность. При такой интерпретации ведущим фактором роста специальной выносливости становится анаэробный гликолиз.
Вывод
Литература
В. В. Шиян Совершенствование специальной выносливости. — М.: ФОН, 1997. — 166 с.
B. Saltin et al. Onset of exercise //Simposium. — Toulouse. — 1972. — P.
Биохимия клетки (энергетика)
В. Н. Селуянов, В. А. Рыбаков, М. П. Шестаков
Глава 1. Модели систем организма
1.1.3. Биохимия клетки (энергетика)
Процессы мышечного сокращения, передачи нервного импульса, синтеза белка и др. идут с затратами энергии. В клетках энергия используется только в виде АТФ. Освобождение энергии, заключенной в АТФ, осуществляется благодаря ферменту АТФ азе, который имеется во всех местах клетки, где требуется энергия. По мере освобождения энергии образуются молекулы АДФ, Ф, Н. Ресинтез АТФ осуществляется в основном за счет запаса КрФ. Когда КрФ отдает свою энергию для ресинтеза АТФ, то образуется Кр и Ф. Эти молекулы распространяются по цитоплазме и активизируют ферментативную активность, связанную с синтезом АТФ. Существуют два основных пути образования АТФ: анаэробный и аэробный (Аулик И. В., 1990; Хочачка П., Сомеро Дж., 1988 и др.).
Анаэробный путь или анаэробный гликолиз связан с ферментативными системами, расположенными на мембране сарко-плазматического ретикулума и в саркоплазме. При появлении рядом с этими ферментами Кр и Ф запускается цепь химических реакций, в ходе которых гликоген или глюкоза распадаются до пирувата с образованием молекул АТФ. Молекулы АТФ тут же отдают свою энергию для ресинтеза КрФ, а АДФ и Ф вновь используются в гликолизе для образования новой молекулы АТФ. Пируват имеет две возможности для преобразования:
1) Превратиться в Ацетил коэнзим А, подвергнуться в митохондриях окислительному фосфорилированию до образования углекислого газа, воды и молекул АТФ. Этот метаболический путь — гликоген-пируват-митохондрия-углекислый газ и вода — называют аэробным гликолизом.

2) С помощью фермента ЛДГ М (лактат-дегидрогеназы мышечного типа) пируват превращается в лактат. Этот метаболический путь — гликоген-пируват-лактат — называется анаэробным гликолизом и сопровождается образованием и накоплением ионов водорода.
Аэробный путь, или окислительное фосфорилирование, связан с митохондриальной системой. При появлении рядом с митохондриями Кр и Ф с помощью митохондриальной КФК азы выполняется ресинтез КрФ за счет АТФ, образовавшейся в митохондрии. АДФ и Ф поступают обратно в митохондрию для образования новой молекулы АТФ. Для синтеза АТФ имеется два метаболических пути:
- 1) аэробный гликолиз;
2) окисление липидов (жиров).
Аэробные процессы связаны с поглощением ионов водорода, а в медленных мышечных волокнах (МВ сердца и диафрагмы) преобладает фермент ЛДГ Н (лактат дегидрогеназа сердечного типа), который более интенсивно превращает лактат в пируват. Поэтому при функционировании медленных мышечных волокон (ММВ) идет быстрое устранение лактата и ионов водорода.
Увеличение в МВ лактата и Н приводит к ингибированию окисления жиров, а интенсивное окисление жиров приводит к накоплению в клетке цитрата, а он угнетает ферменты гликолиза.
Энергия для мышечных сокращений!
Движение любого сочленения осуществляется благодаря сокращениям скелетных мышц. На следующей диаграмме представлен метаболизм энергии в мышце.
Сократительная функция всех типов мышц обусловлена превращением в мышечных волокнах химической энергии определённых биохимических процессов в механическую работу. Гидролиз аденозинтрифосфата (АТФ) как раз и обеспечивает мышцу этой энергией.
Связан с возможностью выполнения работы за счет окисления энергетических субстратов, в качестве которых могут использоваться углеводы, жиры, белки при одновременном увеличении доставки и утилизации кислорода в работающих мышцах.
Для пополнения срочных и кратковременных энергетических запасов и выполнения длительной работы мышечная клетка использует так называемые долговременные источники энергии. К ним относятся глюкоза и другие моносахара, аминокислоты, жирные кислоты, глицеролкомпоненты продуктов питания, доставляемые в мышечную клетку через капиллярную сеть и участвующие в окислительном метаболизме. Эти источники энергии генерируют образование АТФ путем сочетания утилизации кислорода с окислением носителей водорода в электронтранспортной системе митохондрии.
По мере увеличения длительности физических нагрузок происходит постепенное снижение концентрации инсулина в крови. Этот гормон активно участвует в регуляции жирового обмена и при высоких концентрациях тормозит активность липаз. Снижение концентрации инсулина во время длительных физических нагрузок приводит к повышению активности инсулин зависимых ферментных систем, что проявляется в усилении процесса липолиза и увеличении освобождения жирных кислот из депо.
Важность этого регуляторного механизма становится очевидной, когда спортсмены допускают наиболее распространенную ошибку. Нередко, стараясь обеспечить организм легкоусвояемыми источниками энергии, за час до начала соревнований или тренировок они принимают богатую углеводами пищу или концентрированный, содержащий глюкозу, напиток. Такое насыщение организма легкоусвояемыми углеводами приводит через 15—20 минут к повышению уровня глюкозы в крови, а это, в свою очередь, вызывает усиленное выделение инсулина клетками поджелудочной железы. Повышение концентрации этого гормона в крови приводит к усилению потребления глюкозы в качестве источника энергии для мышечной деятельности. В конечном счете, вместо энергетически более выгодных жирных кислот в организме расходуются углеводы. Так, прием глюкозы за час до старта может существенно повлиять на спортивную работоспособность и снизить выносливость к длительной нагрузке.
Каждый из перечисленных биоэнергетических компонентов физической работоспособности характеризуется критериями мощности, емкости и эффективности (табл. 1).